
På feltarbeid i Darwins ånd på Finnmarksvidda
Skrevet av Kim Præbel, Genetikkgruppa, Norges Fiskerihøgskole, UiT Norges Arktiske Universitet
Nå er deler av genetikkgruppen vår på feltarbeid i Finnmark for å samle prøver og data til forskning på en av våre favoritt fisker – nemlig siken (Coregonus lavaretus L.). Vi vil i de neste dagene blogge om hvorfor vi forsker på sik og vise frem de fine omgivelsene vi jobber i. Men først litt bakgrunn om prosjektet.
Siken i Finnmark

Siken som lever i innsjøene i Finnmark er unik sammenlignet med sik fra andre steder i Norge, siden det etter siste istid (5000-9000 år) har blitt dannet minst to nye former (arter) av sik ut i fra den opprinnelig siken som koloniserte innsjøene. Disse tre sikformene lever i dag sammen, men i finnes i ulike deler av innsjøene, og har ulikt levesett (har ulike nisjer).
Vi har også vist at de tre sikformene allerede er blitt genetisk forskjellige hvilket tilsier at denne evolusjonære prosessen er et meget godt eksempel på pågående dannelse av nye arter. Siden denne prosessen foregår i mange innsjøer spredt i hele Finnmark, kan vi bruke hver av innsjøene som et eget replikat når det gjelder vår forskning på artsdannelse. Finnmarksiken og Finnmarksvidda gir oss en unik mulighet til å «se prosessene i et naturlige laboratorium».
Artsdannelse i Finnmark
Vi har sammen med gruppen for Ferskvannsøkologi (UiT) og Kjartan Østbye (HiHM) brukt mange år på å studere Finnmarkssiken for å forstå hvorfor de dannes og hvilke økologiske og miljømessige faktorer som medvirker i denne artsdannelsesprosessen.
Gjennom årene har vi besøkt og undersøkt mer enn 50 innsjøer fra Pasvik-, Tana- og Alta-Kautokeinovassdraget. Disse undersøkelsene har for eksempel vist at antallet av gjellegitterstaver på gjellebuene er forskjellig hos de ulike sikformene, og at antallet av gjellegitterstaver er koblet til hva som siken spiser.
Vi kan derfor si at gjellegitterstavene er en adaptiv karakter som favoriseres i de ulike nisjene når det gjelder de forskjellige sikformene. Vi har så langt identifisert en dyp-levende sik som har 13-22 gjellegitterstaver og som primært spiser dansemygglarver, den vanlige bunnsiken som har 22-32 gjellegitterstaver og som spiser insektlarver, snegler og andre vannlevende invertebrater langs land, og en planktonsik som har mer en 32 gjellegitterstaver som spiser dyreplankton i de frie vannmassene.

Hva har Darwins Galapagos finker og siken i Finnmark til felles?
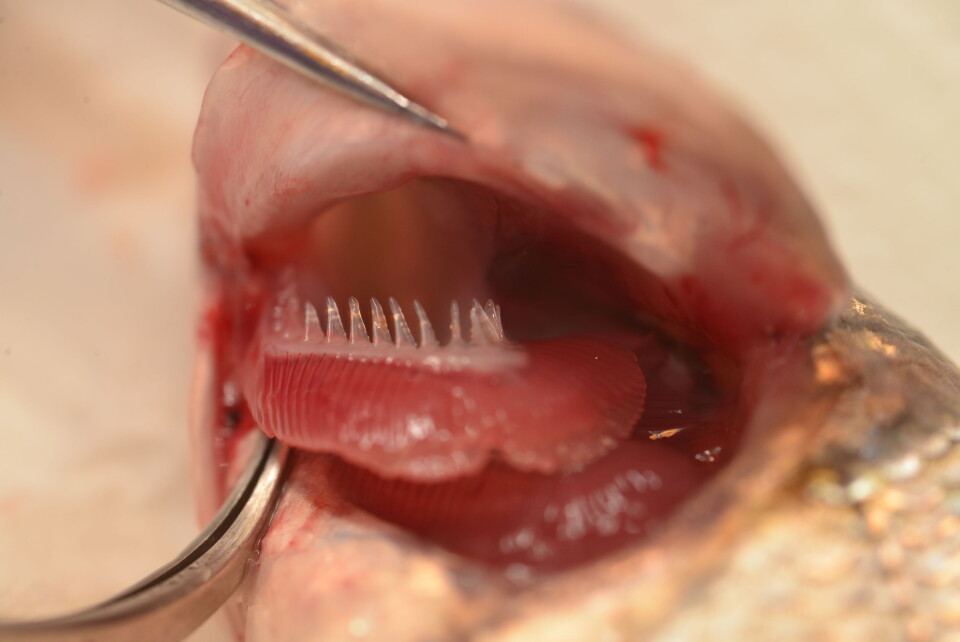
Vi har vist at antall gjellegitterstaver hos de ulike sikformene er koblet til fødevalg og livsstil og at denne prosessen vesentlig er drevet av naturlig seleksjon. Dette betyr at individer som ikke har rett antall gjellegitterstaver i forhold til tilgjengelige byttedyr i en del av innsjøen vil klare seg dårligere enn dem som har rett antall gjellegitterstaver for den nisjen.
Dette kan i ytterste konsekvens bety at slike ”mistilpassede” individer ikke klarer å formere seg og dermed ikke vil gi genene sine videre til den neste generasjonen av sik. Derfor blir sik som har et antall gjellegitterstaver som egner seg best til å innta byttedyrene i en gitt del av innsjøen, for eksempel å filtrerer dyreplankton i de frie vannmassene, favorisert av evolusjonen og det vil over tid dannes populasjoner som er spesialister til å spise ulike byttedyr.
En fin analogi til dette er Darwins Galapagos finker, hvor det er utviklet flere nye former av finker som har nebb spesialisert til å spise ulike typer av frø. Vi tenker derfor at gjellegitterstavene til siken i Finnmark er fiskens nebb.
#Finnmarkfieldwork2016

På dette feltarbeidet fokuserer vi (Masterstudent Julie Bitz-Thorsen, PhD student Katja Häkli, PhD Shripathi Bhat, og jeg) på å samle inn data for forstå hva som ligger bak den genetiske adskillelsen av sikformene, slik at vi kan oppnå en dypere forståelse for hva som driver artsdannelsesprosessen.
Genetiske forskjeller mellom populasjoner oppstår som følge av reproduktiv adskillelse og kan ha mange årsaker. Men for populasjoner som er i fysisk kontakt, og er i et tidlig stadium av reproduktiv adskillelse, er det ofte adskillelse i gytetidspunkt eller plass, eller bevist valg av partner (positiv assortativ parring), som er de drivende faktorene. Siden det ikke påvist assortativt partnervalg hos sik, fokuserer vi på å undersøke om det er adskillelse i gytetidspunkt eller gyteplass som driver de genetiske forskjellene mellom sikformene.
Dette høres kanskje ut som en simpel oppgave, men vi har erfart at det ikke er enkelt. Gytetidspunktet for sik er fra oktober til januar alt etter sikform og innsjø. Innsjøene fryser også over i samme periode og vi skal relativt langt ut på vinteren før enn isen er tykk nok til det er trygt å ferdes på den. Vi kan altså verken fiske fra båt eller fra is for å skaffe materialet vi trenger.
I de siste to årene har vi derfor gamblet med når innsjøene fryser over og planlagt feltarbeidet så tett på islegging som mulig. Sikformene er da så tett på gytingen at vi kan bestemme om det er forskjeller i gytetidspunkt utfra størrelsen på kjønnsorganene og modningsgraden av egg og sperm (mer om dette senere).

Vi tar også prøver til våre genetiske undersøkelser som blant annet har til formål å forstå hvilke biologiske prosesser som påvirkes av naturlig seleksjon i pågående artsdannelse. PhD-student Katja Häkli, Genetikkgruppen, innsamler også prøver til hennes forskning på hvordan adaptive karakterer, som for eksempel gjellegitterstavene, blir uttrykt fra ulike genomiske bakgrunner.
Slik viten vil gi os en dypere innsikt i hvordan miljøet og økologien kan forme utseendet til fisk (og dyr generelt) i en artsdannelsesprosess og også bidra til vår forståelse av hvordan gener og biologiske prosesser responderer på forandringer i miljøet.
Vi har nå satt våre sikgarn og vi vil skrive mer om feltarbeidet og vår forskning på sik i de kommende blogginnlegg.

Referanser og videre lesning
Harrod, C., J. Mallela, and K. K. Kahilainen. 2010. Phenotype-environment correlations in a putative whitefish adaptive radiation. J. Anim. Ecol. 79:1057-1068.
Kahilainen, K., and K. Østbye. 2006. Morphological differentiation and resource polymorphism in three sympatric whitefish Coregonus lavaretus (L.) forms in a subarctic lake. J. Fish Biol. 68:63–79.
Præbel, K., R. Knudsen, A. Siwertsson, K. K. Kahilainen, et al. 2013. Ecological speciation in postglacial European whitefish: rapid adaptive radiations into the littoral, pelagic and profundal lake habitats. Ecology and Evolution 3:4970-4986
Siwertsson, A., R. Knudsen, K. K. Kahilainen, K. Præbel, et al. 2010. Sympatric diversification as influenced by ecological opportunity and historical contingency in a young species lineage of whitefish. Evol. Ecol. Res. 12:929–947.